Acest articol sau această secțiune are bibliografia incompletă sau inexistentă. Puteți contribui prin adăugarea de referințe în vederea susținerii bibliografice a afirmațiilor pe care le conține.
Structura insulinei. Hormon pancreatic secretat de celulele Langerhans, cu rol de reglare a nivelului glucozei în sânge. În stînga este redat monomerul insulinei, cela care se crede ca este activ biologic. Carbonul verde, hidrogenul alb, oxigenul roșu, iar azotul albastru. Partea dreaptă reprezintă hexamerul, forma de stocare. Unitatea monomerului este formată din lanțul A (albastru) și lanțul B cian. Galbenul reprezintă punțile disulfurice, iar sferele violet, ionii de zinc.
Proteinele sunt compuși organicimacromoleculari formați din lanțuri simple sau complexe polipeptidice, având ca unități resturi de aminoacizi. Proteinele prezintă o gamă largă de funcții în organismele vii, precum catalizarea reacțiilor metabolice (prin intermediul enzimelor și complexelor enzimatice), replicarea ADN, semnalizarea celulară, bază structurală în celulă, cât și transportul moleculelor. Diferența majoră dintre proteine este secvența de aminoacizi din catenă, care este dictată cu exactitate de secvența de nucleotide dintr-o anumită genă. Ca urmare a secvenței și a altor proprietăți, proteinele se aranjează spațial, având și o structură tridimensională specifică, necesară pentru activitatea lor. Proteinele sunt prezente în celulele tuturor organismelor vii în proporție de peste 50% din greutatea uscată.
Prima menționare a cuvântului proteină a fost făcută de către Jakob Berzelius, descoperitorul acestora, în scrisoarea sa către Gerhardus Johannes Mulder din 10 iulie1838, scrisoare în care menționează:
"The name protein that I propose for the organic oxide of fibrin and albumin, I wanted to derive from [the Greek word] πρωτειος, because it appears to be the primitive or principal substance of animal nutrition".
(Numele de proteină pe care îl propun pentru denumirea compusului organic rezultat prin oxidarea fibrinei sau albuminei, l-am derivat din grecescul πρωτειος (proteios) deoarece pare a fi substanța primitivă sau principală din nutriția animalelor).
La nivelul ribozomilor se realizează translația informației genetice de la ARNm în lanțul polipeptidic. În nucleu ARN m copiază informația genetică de la ADN nuclear iar apoi migrează în citoplasmă. Fiecare moleculă de ARNt posedă la mijlocul lanțului său un anticodon a căror 3 baze consecutive se pot asocia prin legături de hidrogen cu 3 baze complementare de un codon ARNm [1]. Pe baza acestui anticodon aminoacizii vor fi poziționați conform mesajului genetic din ARNm. Această poziționare a aminoacil-ARNt se realizează la suprafața ribozomului la nivelul a 2 situsuri stereospecifice: situl "P" (peptidil) și situl "A" (aminoacil), situate la suprafața subunităților [2].
Biosinteza proteinelor este un proces prin care fiecare celulă își sintetizează proteinele proprii, prin intermediul unui proces care include multe etape, sinteza începând cu procesul de transcripție și terminând cu procesul de translație. Procesul deși similar, este diferit în funcție de celulă: eucariotă sau procariotă.
Procesul de transcripție necesită prezența unei singure molecule de ADN dublu catenar, numit ADN „șablon”, moleculă care intră în procesul de „inițiere”. Aici acționează enzima ARN polimeraza, enzimă care se leagă de o anumită regiune din molecula de ADN, regiune (denumită promoter), de unde va începe transcripția. Pe măsură ce ARN polimeraza se leagă de promoter, lanțurile de ADN vor începe sa se desfacă. Următorul proces în care intră ADN este procesul de elongație (alungire a catenei). Pe măsură ce ARN polimeraza se mișcă de-a lungul catenei de ADN, are loc sinteza ribonucleotidelor complementare (ARNm - ARN mesager). Acest ARN, după cum îi arată și numele, se poate deplasa și în alte părți ale celulei cum ar fi reticulul endoplasmatic sau citoplasma.
Gruparea 5' se găsește în capătul 5' final al moleculei de ARNm, și este format din guanosină grefată printr-o legătură de tip 5'- 5' de molecula de ARN prin intermediul unei legături trifosfat.
Are loc adiția unei grupări 5', grupare dinucleotidică care are rolul de a asigura stabilitatea ARN și de a-l transforma în ARN matur. O secvență de aminoacizi este grefată în poziția 3' terminală pentru protecție dar și pentru a sluji drept șablon pentru procesele următoare.Mai departe are loc formarea ARN, care este apoi utilizat in ribozomi pentru sinteza proteinelor. La procariote legarea ARN de ribosomi are loc după ce acesta este îndepărtat de nucleoid; în contrast la eucariote acest proces are loc chiar în membrana nucleară și apoi translocat în citoplasmă. Rata sintezei proteice poate ajunge la circa 20 aminoacizi la procariote, mult mai puțin la eucariote.
În timpul translației ARNm transcris din ADN este decodat de ribozomi pentru sinteza proteinelor.Acest proces este divizat în 3 etape:
Inițierea
Elongarea
Faza terminală.
Ribozomul are situsuri de legare care permit altei molecule de ARNt (ARN de transfer), să se lege de o moleculă de ARNm, proces însoțit de prezența unui anticodon. Pe măsură ce ribozomul migrează de-a lungul moleculei de ARNm (un codon o dată) o altă moleculă de ARNt este atașată ARNm. Are loc eliberarea ARNt primar, iar aminoacidul care este atașat de acesta este legat de ARNt secundar, care îl leagă de o altă moleculă de aminoacid. Translația continuă pe măsură ce lanțul de aminoacid este format. La un moment dat apare un codon de stop, o secvență formată din 3 nucleotide (UAG, UAA), care semnalează sfîrșitul lanțului proteic. Chiar după terminarea translației lanțurile proteice pot suferi modificări post-translaționale și plierea lanțului proteic, responsabilă de structura secundară și cea terțiară. Modificările post-translaționale se referă la posibilitatea formării de legături disulfidice, sau de atașarea la scheletul proteic a diferite grupări ca rol biochimic: acetat, fosfat etc.
Procesul de sinteză chimică poate avea loc în laborator, dar pentru lanțuri mici de proteine. O serie de reacții chimice cunoscute sub denumirea de sinteza peptidelor, permit producerea de cantități mari de proteine. Prin sinteza chimică se permite introducerea în lanțul proteic a aminoacizilor ne-naturali, atașarea de exemplu a unor grupări fluorescente. Metodele sunt utilizate în biochimie și in biologia celulei. Sinteza are la bază cuplarea grupării carboxil -COOH (carbon terminus) cu gruparea -amino -NH2 (segmentul N terminus).
Se cunosc 2 metode de sinteză pe cale chimică_
Sinteza în fază lichidă, metoda clasică, care a fost înlocuită cu sinteza în fază solidă.
Sinteza în fază solidă (Solid-phase peptide synthesis SPPS), a cărei bază a fost pusă de Robert Bruce Merrifield. Prin aceată metodă, se pot sintetiza proteine D, cu aminoacizi D. În prima fază Merrifield a folosit metoda tBoc (terț-butil-oxi-carbonil). Pentru înlăturarea acestuia din lanțul peptidic se folosește acidul fluorhidric (HF), care este foarte nociv, periculos, iar din acest motiv, metoda nu se mai utilizează. Atunci cînd este vorba de sinteza analogilor peptidici non-naturali de tip bază (depsi-peptidele) este necesară.
O altă metodă este cea introdusă de R.C. Sheppard în anul 1971, și are la bază folosirea Fmoc (fluorenil metoxi carbonil), iar pentru îndepărtarea acesteia se folosește de obicei mediu bazic asigurat de o soluție 20% piperidină/DMF (dimetil formamidă). Îndepărtarea grupării din lanțul proteic se face prin incubare în acid trifluoracetic (TFA).
Această grupă de protejare cu simetrie ortogonală se folosește în multe sinteze chimice.
Protejarea grupării prin intermediul Fmoc este de obicei lentă, deoarece anionul nitro produs la sfîrșitul reacției nu este un produs favorabil desfășurării reacției.
Datorită compoziției, fiind formate exclusiv din aminoacizi, se întâlnesc alături de alți compuși importanți de tipul polizaharidelor, lipidelor și acizilornucleici începând cu structura virusurilor, a organismelor procariote, eucariote și terminând cu omul. Practic nu se concepe viață fără proteine. Proteinele pot fi enzime care catalizează diferite reacții biochimice în organism, altele pot juca un rol important în menținerea integrității celulare (proteinele din peretele celular), în răspunsul imun și autoimun al organismului.
Majoritatea microorganismelor și plantelor pot sintetiza toți cei 20 aminoacizi standard, în timp ce organismele animale obțin anumiți aminoacizi din dietă (aminoacizii esențiali). Enzime cheie, cum ar fi de exemplu aspartat kinaza, enzimă care catalizează prima etapă în sinteza aminoacizilor lisină, metionină și treonină din acidul aspartic, nu sunt prezente în organismele de tip animal. La aceste organisme aminoacizii se obțin prin consumul hranei conținând proteine. Proteinele ingerate sunt supuse acțiunii acidului clorhidric din stomac și acțiunii enzimelor numite proteaze, proces în urma căruia lanțurile proteice sunt scindate (denaturate). Ingestia aminoacizilor esențiali este foarte importantă pentru sănătatea organismului, deoarece fără acești aminoacizi nu se poate desfășura sinteza proteinelor necesare organismului. De asemenea, aminoacizii sunt o sursă importantă de azot; unii aminoacizi nu sunt utilizați direct în sinteza proteică, ci sunt introduși în procesul de gluconeogeneză, proces prin care organismul asigură necesarul de glucoză în perioadele de înfometare (mai ales proteienele aflate în mușchi).
În funcție de compoziția lor chimică ele pot fi clasificate în:
Holoproteine cu următoarele clase de proteine
Proteine globulare (sferoproteine) sunt de regulă substanțe solubile în apă sau în soluții saline: protaminele, histonele, prolaminele, gluteinele, globulinele, albuminele.
Proteinele fibrilare (scleroproteinele) caracteristice regnului animal, cu rol de susținere, protecție și rezistență mecanică: colagenul, cheratina și elastina.
Heteroproteinele sunt proteine complexe care sunt constituite din o parte proteică și o parte prostetică; în funcție de această grupare se pot clasifica astfel:
Datorită formării aproape în exclusivitate din aminoacizi, putem considera proteinele ca fiind de fapt niște polipeptide, cu masă moleculară foarte mare, între 10.000 și 60.000.000. Masa moleculară se determină prin diferite metode, mai ales în cazul proteinelor cu masa moleculară foarte mare ca de exemplu proteina C reactivă.
Masa moleculară a diferitelor proteine
Denumirea proteinei
Sursa proteinei/Izolată din
Masa moleculară
Lactalbumină
lapte
17.000
Gliadina
grâu
27.500
Insulina
pancreas
12,000
Hordeina
orz
27.500
Hemoglobina
globule roșii
68.000
Hemocianina
moluște(sânge) , artropode(sânge)
2.800.00
Miozina
mușchi
850.000
Pepsină
stomac
36.000
Peroxidaza
rinichi
44.000
Virusul mozaicului tutunului (capsida)
tutun
17.000.000
Deoarece la multe proteine masa moleculară apare ca un multiplu de 17,500, multă vreme s-a mers pe ipoteza că particulele proteice sunt formate prin unirea mai multor molecule de bază ce au masa moleculară în jurul valorii de 17,500. Aceste molecule de bază s-ar putea uni între ele prin așa numitele valențe reziduale, ducând la formarea de agregate moleculare. Atunci când are loc ruperea acestor valențe reziduale ar avea loc doar modificarea proprietăților fizice ale proteinelor, în timp ce dacă are loc ruperea legăturilor principale (legăturile peptidice), proteina își modifică proprietățile fizico-chimice.
Proteinele sunt substanțe solide, macromoleculare, solubile în general în apă și insolubile în solvenți organici nepolari. Unele proteine sunt solubile în apă dar insolubile în alcool, altele sunt solubile în soluții apoase de electroliți, acizi organici. Datorită gradului diferit de solubilitate în diferiți solvenți, proteinele se pot izola, identifica și separa. Solubilitatea lor depinde foarte mult de legăturile care se stabilesc între grupările libere de la suprafața macromoleculelor și moleculele solventului. La suprafața macromoleculelor proteice se găsesc grupări libere de tip polar,-COOH, -NH2, -OH, -SH, -NH, grupări cu caracter hidrofil care favorizează dizolvarea proteinelor în apă. De asemenea există grupări de tip apolar, hidrofobe, de regulă radicali de hidrocarburi -CH3, -C6H5, -C2H5, care favorizează dizolvarea proteinelor în alcool. Însă în marea lor majoritate predomină grupările polare, determinante pentru caracterul hidrofil. În contact cu apa proteinele greu solubile manifestă fenomenul de gonflare, datorită tendinței de hidratare datorată grupărilor polare. Gelatina de exemplu se îmbibă foarte puternic cu apa dînd naștere prin răcire la geluri. La dizolvarea proteinelor în apă, are loc fenomenul de formare a coloizilor hidrofili. S-a constatat că în soluții diluate se găsesc macromolecule proteice izolate, iar în cazul soluțiilor concentrate se formează agregate de macromolecule proteice. Soluțiile coloidale ale proteinelor, coagulează prin încălzire, prezintă efectul Tyndall (dispersia fasciculului de lumină).
Proteinele, la fel ca și aminoacizii, sunt substanțe amfotere și formează în soluții apoase amfioni:
, în prezența H2O
În mediu acid proteinele se comportă ca baze slabe, ele primind protoni și formând cationi proteici:
, cation al proteinei. Reacția stă la baza electroforezei proteinelor, datorită incărcării pozitive cationii migrează spre catod, fenomen numit cataforeză, proteina fiind în acest caz electropozitivă.
În mediu bazic proteinele se comportă ca acizii slabi, ele cedând protoni, se formează astfel anioni proteici, care migrează spre anod fenomenul fiind denumit anaforeză, proteina avînd încărcare electronegativă.
, anion al proteinei.
Datorită caracterului amfoter proteinele pot neutraliza cantități mici de substanță acidă sau bazică, avind în acest fel rol de soluție tampon, prin acest lucru contribuind la menținerea echilibrului acido-bazic al organismului. În general caracterul amfoter este imprimat de grupările -NH2 și -COOH libere care nu sunt implicate în legăturile peptidice. Dacă în molecula proteinei există mai mulți aminoacizi dicarboxilici atunci molecula se va comporta ca un acid slab, iar în cele în care predomină aminoacizii diaminați se comportă ca baze slabe. Chiar dacă într-o moleculă există un număr egal de grupări amino si carboxil, deci teoretic molecula ar trebui sa fie neutră, în realitate datorită gradului de ionizare mult mai mare a grupării carboxil față de gruparea amino, molecula proteinei va avea un caracter slab acid, în soluția ei întâlnindu-se amfiioni proteici, anioni proteici și protoni (H+ ).
Prin acidulare echilibrul reacției se deplasează spre formarea de cationi proteici. La o anumită concentrație a H+, proteina devine neutră deoarece gruparea aminică și cea carboxilică sunt la fel de disociate și deci molecula este neutră din punct de vedere electric. În acel moment se vor găsi în soluție amfiioni, H+, ioni hidroxil -HO; pH-ul la care soluția unei proteine conține anioni și cationi în proporție egală poarta denumirea de punct izoelectric, se notează cu pHi, fiind o constantă foarte importantă a proteinelor. Fiecare proteină la punctul izoelectric are un comportament specific, avînd o solubilitate si reactivitate chimică minimă; de asemenea hidratarea particulelor coloidale, vâscozitatea și presiunea osmotică sunt de asemenea minime. Precipitarea proteinei la punctul izoelectric este în schimb maximă, dar nu se deplasează sub influența curentului electric. De obicei valorile punctului izoelectric variază între 2,9 și 12,5 [1][2]
și se determină prin diferite metode: potențiometrice, electroforetice.
Sub acțiunea diferiților factori fizici (ultrasunete, radiații cu diferite lungimi de undă, căldură), factori chimici (acizi, baze, diferiți solvenți organici), sau mecanici (agitare), are loc fenomenul de precipitare a proteinelor, precipitarea care poate fi reversibilă sau ireversibilă.
Precipitarea reversibilă se poate produce sub acțiunea soluțiilor concentrate ale sărurilor alcaline dar și în prezența unor dizolvanți organici miscibili cu apa în orice proporție, cum sunt de exemplu acetona și alcoolul. În cadrul acestei precipitări molecula proteinei suferă unele modificări fizico-chimice, dar nu are loc afectarea structurii moleculare. Puterea de precipitare a proteinelor de către diferiți ioni este data de seria liofilă a lui Hofmeister [3]. Dacă anionul rămîne același, puterea de precipitare a cationilor scade în următoarea ordine:
Li+>Na+>NH4+>
cănd cationul ramîne același anionii se comportă astfel:
SO42->PO43->CH3COO->Citrat->tartrat->Cl->NO3->ClO3->Br->I->SCN-.
Solvenții de tipul alcoolului sau acetonei, în funcție de concentrația lor, pot forma fie precipitate reversibile, fie ireversibile. Sărurile alcaline au un comportament diferit față de proteine, în soluții diluate mărind solubilitatea proteinelor, iar în soluții concentrate determinînd precipitarea lor reversibilă. De altfel soluțiile sărurilor alcaline de diferite concentrații se folosesc pentru precipitarea fracționată a proteinelor din amestecuri.
În cursul acestei precipitări molecula proteinei suferă modificări fizico-chimice ireversibile avînd loc și modificarea structurii moleculare. De regulă se produce la adăugarea de soluții ale metalelor grele (Cu,Pb, Hg, Fe), a acizilor minerali tari (HNO3, H2SO4) acidul tricloracetic, a soluțiilor concentrate de alcool sau acetonă, sau, în cazul anumitor proteine, în prezența căldurii. Prin precipitare ireversibilă proteinele își pierd activitatea biologică (enzimatică, hormonală, etc.), are loc o descreștere a solubilității, modificarea activității optice și, de asemenea, sunt mai ușor de degradat sub acțiunea unor enzime proteolitice. Prin îndepărtarea factorilor care au dus la precipitare, proetienele nu revin la forma lor inițială și nu iși pot reface structura moleculară. Proteinele precipitate își pierd din proprietățile hidrofile "obțînînd" proprietăți hidrofobe.
Din punct de vedere chimic, proteinele sunt heteropolimeri constituiți din 20 de L-α aminoacizi (așa numiții aminoacizi standard, vezi tabelul), în care grupările carboxil se pot combina cu grupările amino formând legături peptidice și rezultând lanțurile peptidice.
Aminoacizii standard au proprietăți variate, proprietăți care sunt direct responsabile de structura tridimensională a proteinei, dar și de proprietățile acesteia.
(4)
În lanțul polipeptidic aminoacizii formează legăturile peptidice prin cuplarea grupei carboxil cu o grupă amino; odată legat în lanțul proteic aminoacidul se "transformă" în aminoacid "rezidual" iar atomii de carbon, azot, hidrogen și oxigen implicați în legături formează "scheletul" proteinei. Atunci cînd lanțul proteic se termină cu o grupă carboxil poartă denumirea de carboxi-terminus (sau C -terminus), în timp ce, dacă se termină cu gruparea amino, devine amino-terminus (N-terminus).
Responsabile de proprietățile chimice sunt aceleași grupări carboxil și amino libere, neimplicate în formarea legăturilor peptidice, însă mai intervin și diferiții radicali grefați pe scheletul proteinei.
Datorită grupărilor carboxil și amino libere ele dau aceleași reacții ca și la aminoacizi.
Caracterul amfoter este responsabil de formarea de săruri atât cu bazele cât și cu acizii
Legătura peptidică este responsabilă de formarea de combinații complexe denumite chelați.
Prezența diferiților radicali alchilici, sau arilici determină formarea unor derivați ai substanțelor proteice (derivații halogenați și nitrici sunt cei mai importanți).
Datorită existenței anumitor aminoacizi în molecula proteinelor, a legăturilor peptidice formate în molecula proteinei dar și grupările funcționale libere sunt responsabile de reacțiile de culoare.
Denumirea reacției
Reactivul folosit
Culoarea rezultată
Tipul de aminoacid identificat
Xantoproteică
acid azotic,hidroxid de amoniu
portocalie
aminoacizii aromatici (formează nitroderivați)
Millon
azotat de mercur în acid azotic/azotit
precipitat roșu cărămiziu sau colorație roșie
aminoacizi ciclici cu grupare hidroxil (tirosina)
Sulfurii de plumb
Acetat sau azotat de plumb în mediu alcalin
precipitat negru de sulfură de plumb
aminoacizi cu sulf în moleculă : cisteină, metionină cistină
După cum s-a văzut mai sus lanțurile peptidice sunt formate de grupările carboxil și aminice a aminoacizilor; există de fapt 2 forme pentru fiecare proteină, numite forme de rezonanță:
una datorată dublei legături care asigură rigiditatea și nu permite rotația în jurul axei sale;
a doua formă de rezonanță este dată de unghiul diedru φ(planul atomilor C'-N-Cα-C'), ψ (planul atomilor N-Cα-C'-N), ω (planul atomilor Cα-C'-N-Cα), unghiurile φ și ψ pot avea diferite valori fiind responsabile de gradul de libertate a proteinelor, controlînd structura tridimensională a lanțului proteic.
Structura substanțelor proteice este încă insuficient cunoscută datorită dinamicității structurii proteinelor, deoarece ele sunt în permanență supuse unor procese de sinteză și de degradare. Pentru evidențierea succesiunii aminoacizilor în structura proteinelor se folosesc 2 metode:[4]
Degradarea EdmanPrin degradarea Edman se poate identifica o secvență de pînă la 30 aminoacizi, cu o eficiență de 98%/aminoacid. Un alt avantaj ar fi cantitatea de numai 10-100 picomoli de peptidă necesari pentru determinare.
Degradarea Edman folosește ca reactiv izotiocianatul de fenil care evidențiază selectiv aminoacidul. Grupa amino terminală se adiționează la izotiocianat trecînd printr-un derivat de tiouree. După ce se tratează cu un acid slab, aminoacidul marcat sub formă de feniltiohidantoină se detașează de restul polipeptidei. Aceasta cu noul său aminoacid terminal poate fi supusă la un nou ciclu de tratări pentru identificarea următoarei grupe amino.
Degradarea Sanger are la bază tratarea polipetidei cu fluoro-2,4-dinitrobenzen, avind loc atacul reactivului asupra grupării amino a aminoacidului N-terminal. Metoda Sanger are dezavantajul degradării complete a polipeptidei.
Unghiul legăturii între C1 și N este aproape de 1800, similar cu unghiul valenței din molecula apei
S-a ajuns la concluzia că există 4 niveluri (structuri), care alcătuiesc edificiul proteic.
Structura primară este dată de aminoacizii care intră în lantul proteic prin formarea legăturilor peptidice.
În structura primară se observă lanțul de aminoacizi
În proteinele naturale legătura peptidică se stabilește între gruparea carboxilică de la C1 și gruparea aminică de la C2, încît lanțul peptidic va fi format dintr-o succesiune de unități CO-NH-CH, legate cap-cap.
La unul din capetele lanțului peptidic se găsește o grupare -NH2 liberă, iar la celălat capăt seaflă o grupare -COOH liberă
Legătura peptidică -CO-NH- se găsește în același plan, iar carbonul -CH- se poate roti, putînd să apară în planuri diferite. Datorită lungimii relativ mici a catenelor laterale, ele se pot aranja de o parte și de alta a lanțului proteic, astfel că lanțul proteic nu este ramificat.
Datorită deplasării altrenative a unui electron de la gruparea -NH la C=O se produce oscilarea dublei legături de la atomul de carbon și oxigen la atomul de azot, fomrîndu-se astfel cele 2 forme mezomere.
Datorită numărului relativ mic de aminoacizi care intră în structura proteinelor, teoretic ar trebui să se formeze proteine cu masa moleculară în jur de 4200. Însă în realitate masele moleculare ale proteinelor au valori de peste 10,000 ceea ce a dus la concluzia că cel puțin o parte de aminoacizi se repetă de mai multe ori în cadrul unei molecule. Ipoteza că proteinele sunt formate din lanțuri lineare de aminoacizi a fost fomulată pentru prima dată în anul 1902, la a 74-a reuniune a Societății Oamenilor de Stiință din Germania, ținută în orașul Karlsbad, de către Franz Hofmeister (ținînd cont de reacția biuretului) și Emil Fischer (care aduce clarificări asupra scheletului proteic). Ipoteza că în molecula proteinelor există legături amidice fusese elaborată de chimistul francez E Grimaux încă din anul 1882. În ciuda evidențelor care demonstrau faptul că proteinele supuse acțiunii proteolitice se scindează în oligopeptide, ideea că lanțul proteic este liniar, au fost idei greu de "digerat". În perioada respectivă, numeroși savanți (William Astbury, Hermann Staudinger), punînd la îndoială acest lucru, prin argumentarea că legăturile amidice nu sunt îndeajuns de puternice pentru a susține o moleculă proteică lungă.
Cu timpul au apărut diverse ipoteze:
Ipoteza coloidală care susținea ca proteinele sunt ansambluri moleculare coloidal formate din molecule mai mici - ipoteză contrazisă de măsurarea ultracentrifugării de către Svedberg care arată faptul că proteinele sunt molecule bine definite, au greutate moleculară, iar prin electroforeză Arne Tiselius demonstrează că proteinele sunt molecule unice.
Ipoteza a 2-a, numită ipoteza ciclol, avansată de Dorothy Wrinch, are la bază 3 elemente:
Ciclol reaction în care gruparea carbonil și gruparea amino a 2 peptide se incrucișează C=O + HN → C(OH)-N (așa numita legătură în cruce); aceste legături sunt de tip covalent, similare cu legăturile covalente de hidrogen propuse de William Astbury, pentru a explica stabilitatea structurii proteice.
Lanțurile beta vecine au la bază o serie de reacții de tip ciclol
Structura proteinelor mici corespund așa numitelor "solid de tip Platon", fără ca să existe colțuri libere.
Structura secundară se referă la forma și la lungimea lanțurilor polipeptidice, proprietăți induse de legăturile de hidrogen. Cele mai întîlnite tipuri de structura secundară sunt alpha helixul și lanțurile beta.
Elicea alpha se formează prin rotația unui lanț polipeptidic în jurul propriei axe
Alte helix-uri cum ar fi helixul 310 și helixul π sunt, din punct de vedere energetic, favorabile formării legăturilor de hidrogen, dar sunt rareori observat în proteinele naturale exceptînd părțile terminale ale helixului α în timpul formării scheletului proteic (de obicei centrul helixului). Aminoacizii au un comportament diferit vis-a-vis de posibilitatea formării structurii secundare. Prolina și glicina sunt cunoscuți ca așa numiții "helix breakers" (spărgători de helix), deoarece afectează configurația scheletului proteic; ambii aminoacizi au abilități conformaționale neobișnuite și de regulă se găsesc în colțurile scheletului proteic. Aminoacizii care preferă să adopte conformația helixului proteic fac parte din așa numita serie MALEK (codurile formate din 1 literă a aminoacizilor: metionină, alanină, leucină, acid glutamic și lizina); prin contrast aminoacizii aromatici (triptofanul, tirosina și fenilalanina, dar și aminoacizii cu legare prin carbonul beta (izoleucina, valina și treonina, adoptă configurația β.
Structura secundară cunoaște cîteva ipoteze privind formarea ei:
Teoria polipeptidică formulată de către E. Hoffmeister în 1902 și dezvoltată ulterior de către E.Fischer, are la bază conceptul conform căruia moleculele proteice sunt formate din lanțuri polipeptidice foarte lungi. Teoria are cîteva dezavantaje:
nu explica diferențierea biologică a anumitor proteine
unele proteine sunt rezistente la acțiunea enzimelor proteolitice (deși datorită lungimii lanțului nu ar trebui)
Teoria plierii și răsucirii lanțului polipeptidice a fost elaborată de către Corey și Pauling în 1943 și a fost confirmată prin spectrele de difracție cu raze X, microscopului electronic , prin măsurarea unghiurilor de valență, a distanțelor interatomice, au confirmat faptul că lanțul polipeptidic se găsește sub formă pliată.
Structura în foaie pliantă. Plierea catenei are loc prin formarea legăturilor de hidrogen între gruparea carboxilică a unui aminoacid și gruparea aminică a aminoacidului vecin. Lanțul polipetidic pliat se prezintză ca o panglică îndoită alternativ la dreapta și la stînga, plierea avînd loc în dreptul carbonilor metinici. Mai multe lanțuri pliate polipeptidice pliate dau naștere unei rețele, între aceste lanțuri pliate putîndu-se de asemenea forma legături de hidrogen, acestea fiind în număr mai mare cînd grupările terminale a 2 lanțuri sunt aranjate diferit (-NH2 și COOH, sau HOOC-și -NH2). Catenele polipeptidice pliate predomină în proteinele fibrilare și mai puțin în cele globulare. După valoarea perioadei de identitate se cunosc mai multe tipuri de proteine cu structură pliată. Prin perioada de identitate se înțelege distanța cea mai mică la care se repetă aminoacizii identici din moleculă.
Structura α elicoidală, ipoteză lansată de Corey și Pauling, ipoteză conform căreia lanțul polipeptidic se poate prezenta și înfășurat sub formă de spirală. În acest model, fiecare spiră conține de obicei 27 aminoacizi, iar distanța între spire este de 5,44 A0. Fiecare aminoacid mărește spira cu 1,47 A0. În fața fiecărei grupări -CO- va apare la o distanță de 2,8A0. o grupare NH de la al treilea aminoacid. Între aceste grupări se stabilesc punțile de hidrogen care asigură stabilitatea α helix-ului. În acest model lanțul polipeptidic se prezintă sub forma unui surub cu pasul fie spre dreapta, fie spre stînga. În cazul proteinelor naturale, acestea datorită conținutului în L-aminoacizi, pasul helixului va fi spre dreapta, catenele laterale ies în afara corpului propriu-zis putînd reacționa fie cu moleculele solventului fie cu alte catene polipeptidice. Canalul format în interiorul helixului este foarte îngust, în el nu poate pătrunde molecula solventului. Legăturile peptidice sunt plane, iar 2 planuri consecutive -CO-NH- formează un unghi de 1800, rotirea lanțului se face la carbonul α(metinic).
Prin intermediul cristalografiei cu raze X s-a dovedit faptul că macromoleculele proteice au o conformație tridrimensională, realizată de obicei prin intermediul cuplării mai multor lanțuri polipeptidice scurte între ele, cuplare care duce la formarea fibrelor proteice;legăturile intercatenare pot fi principale sau secundare:
Legături de hidrogen, sunt legături coordinativ heteropolare care se stabilesc cu ușurință între gruparea carbonil C=O (electronegativă) și gruparea NH- (electropozitivă), din 2 lanțuri polipeptidice alăturate, sau în cazul formelor lactam-lactimă între gruparea -OH și azotul iminic =NH
Legăturile de hidrogen au lungimea cuprinsă între 2,7-3,1A și energia de 3-7Kcal/mol la peptide, iar la apă 2-3Kcal/mol
[6].Legăturile de hidrogen se pot stabili și între catenele lateralecare au grupări carboxil, hidroxil, amino sau tiolice. Din punct de vedere energetic legătura de hidrogen nu este puternică dar datorită răspîndirii relativ uniforme de-a lungul scheletului proteic oferă proteinei stabilitatea necesară.
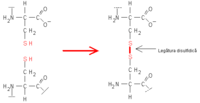
Legături disulfidice Legătura disulfidică este foarte puternică ,50-100kcal/mol și are un rol foarte importantîn stabilizarea arhitecturii spațiale a moleculei proteice. Legătura este rezistentă la hidroliză, însă se poate desface iar prin reducere formează tioli(SH), iar prin oxidare formează acizi. În general legătura sulfidică se întîlnește la proteinele transformate, care au o rezistență mecanică mare.
În afară de aceste legături se mai pot stabili alte tipuri de legături:
legături ionice (stabilite de obicei între grupările aminice și cele carboxilice ionizate), legături de tip van der Waals (legături electrostatice slabe care se stabilesc între radicalii hidrofobi), legături fosfodiesterice (între 2 resturi de serină și acid fosforic), legături eterice (stabilite la nivelul aminoacizilor cu grupări hidroxilice).
Structura cuaternară se referă la modul în care se unesc subunitățile proteice. Enzimele care catalizează asamblarea acestor subunități poartă denumirea de holoenzime, în care o parte poartă denumirea de subunități reglatoare și subunități catalitice.
Vedere 3 D a hemoglobinei cele 4 subunități roșu și galben, iar unitatea hemică verde.numele de hemoglobină vine este formată din hem și globină, denumire ce denotă faptul că hemoglobina are la bază proteine globulare cuplate cu o grupare hem
. Proteine care au structura cuaternară :hemoglobina, ADN polimeraza și canalele ionice, dar și nucleozomi și nanotubuli, care sunt complexe multiproteice.Fragmentele proteice pot suferi transformări în structura cuaternară, transformări care se reflectă fie în structurile individuale fie în reorientările fiecărei subunități proteice. Numărulsubunităților din oligomerice sunt denumite prin adăugarea sufix-ului -mer (grecescul pentru subunitate), precedat de numele subunității.





















{kind=link}
{kind=link}
{kind=link}
{kind=link}